Молекула РНК также полимер, мономерами которого является рибонуклеотиды, РНК представляет собой одноцепочную молекулу. Она построена таким же образом, как и одна из цепей ДНК. Нуклеотиды РНК похожи на нуклеотиды ДНК, хотя и не тождественны им. Их тоже четыре, и они состоят из осатков азотистого основания, пентозы и фосфорной кислоты. Три азотистых основания совершенно такие же, как в ДНК: А , Г и Ц . Однако вместо Т у ДНК в РНК присутствует близкое по строение пиримидиновое основание – урацил (У ). основное различие между ДНК и РНК – это характер углевода: в нуклотидах ДНК моносахарид – дезоксирибоза, а в РНК – рибоза. Связь между нуклеотидами осуществляется, как и в ДНК, через сахар и остаток фосфорной кислоты. В отличие от ДНК, содержание которой в клетках определенных организмов постоянно, содержание РНК в них колеблется. Оно заметно выше там, где происходит интенсивный синтез.
В отношении выполняемых функций различают несколько видов РНК.
Транспортная РНК (тРНК). Молекулы тРНк самые короткие: они состоят всего из 80-100 нуклеотидов. Молекулярная масса таких частиц равна 25-30 тыс. Транспортные РНК в основном содержатся в цитоплазме клетки. Функция их состоит в переносе аминокислот в рибосомы, к месту синтеза белка. Из общего содержания РНК клетк на долю тРНК приходится около 10%.
Рибосомная РНК (рРНК). Это крупные молекулы: в их состав входит 3-5 тыс. нуклеотидов, соотвественно их молекулярная масса достигает 1-1,5 млн. Рибосомные РНК составляют существенную часть рибосомы. Из общего содержания РНК в клетке на долю рРНК приходится около 90%.
Информационная РНК (иРНК), или матричная РНК (мРНК), содержится в ядре и цитоплазме. Функция ее состоит в переносе информации о структуре белка от ДНК к месту синтеза белка в рибосомах. На долю иРНК приходится примерно 0,5-1% от общего содержания РНК клетки. Размер иРНК колеблется в широких пределах – от 100 до 10000 нуклеотидов.
Все виды РНК синтезируются на ДНК, которая служитсвоего рода матрицей.
ДНК – носитель наследсвенной нформации.
Каждый белок представлен одной или несколькими полипиптидными цепями. Участок ДНК, несущий информацию об одной полипиптидной цепи, называют геном . Совокупность молекул ДНК клетки выполняет функцию носителя генетической информации. Генетическая информация передается как от материнской клетки дочерним клеткам, так и от родителей детям. Ген является единицей генетической , или наследственной, информации.
ДНК – носитель генетической информаци в клетке – непосредственного участия в синтезе белков не принимает. В клетках эукариот молекулы ДНК содержатся в хромосомах ядра и отделены ядерной оболочкой от цитоплазмы, где происходит синтез белков. К рибосомам – местам сборки белков – высылается из ядра несущий информацию посредник, способный пройти через поры ядерной оболочки. Таким посредником является информационная РНК (иРНК). По принципу комплементарности она синтезируется на ДНК при участие фермента, называемого РНК-полимеразой .

Информационная РНК – это однонитевая молекула, и транскрипция идет с одной цепи двунитевой молекулы ДНК. Она является копией не всей молекулы ДНК, а только части ее – одного гена у эукариот или группы рядом расположенных генов, несущих информацию о структуре белков, необходимых для выполнения одной функции, у прокариот. Такую группу генов называют опероном . В начале каждого оперона находится своего рода посадочная площадка для РНК-полимеразы, называемая промотором .это специфическая последовательность нуклеотидов ДНК, которую фермент «узнает» благодаря химическому сродству. Только присоединившись к промотору, РНК-полиммераза способна начать интез РНК. Доядя до конца оперона, фермент встречает сигнал (в виде определенной последоватльности нуклеотидов), означающий конец считывания. Готовая иРНК отходит от ДНК и направляется к месту синтеза белков.
В процессе транскрипции можно выделить четыре стадии: 1) связывание РНК -полимеразы с промотором; 2) инициация – начало синтеза. Оназаключается в образовании первой фосфодиэфирной связи между АТФ или ГТФ и вторым нуклеотидом синтезирующейся молекулы РНК; 3) элонгация – рост цепи РНК; т.е. последовательное присоединение нуклеотидов друг к другу в том порядке, в котором стоят комплементарные им нуклеотиды в транскрибируемой нити ДНК. Скорость элонгации 50 нуклеотидов в секунду; 4) терминация – завершение синтеза РНК.
Пройдя через поры ядерной оболочки, иРНК направляется к рибосомам, где осуществляется расшифровка генетической информации – перевод ее с «языка» нуклеотидов на «язык» аминокислот. Синтез полипептидных цепей по матрице иРНК, происходящий в рибосомах, называют трансляцией (лат. translation – перевод).
Аминокислоты, из котрых синтезируютсябелки, доставляются к рибосомам с помощью специальных РНК, называемых транспортными (тРНК). В клетке имеется столько же разных тРНК, сколько кодонов, шифрующих аминокислоты. На вершине «листа» каждой тРНК имеется последовательность трех нуклеотидов, комплементарных нуклеотидам кодона в иРНК. Ее называют антикодоном. Специальный фермент – кодаза – опознает тРНК и присоединяет к «черешку листа» аминокислоту – только ту, которая кодируется триплетом, комплементарным антикодону. На образование ковалентной связи между тРНК и «своей» аминокислотой затрачивается энергия одной молекулы АТФ.
 Для
того чтобы аминокислота включилась в
полипептидную цепь, она должна оторваться
от тРНК. Это становится возможным, когда
тРНК поступает на рибососму и антикодон
узнает свой кодон в иРНК. В рибосоме
имеется два участка для связывания двух
молекул тРНК. В один из этих участков,
называемый акцепторным
,
поступает тРНК с аминокислотой и
присоединяется к своему кодону (I).
Эта аминокислота присоединяет к себе
(акцептирует) растущую цепь белка (II)?
Между ними образуется пептидная связь.
тРНК, к которой теперь присоединяется
вместе с кодоном иРНК в донорный
участок рибосомы. В освободившийся
акцепторный участок приходит новая
тРНК, связанная с аминокислотой, которая
шифруется очередным кодоном (III).
Из донорного участка сюда вновь
переносится оторвавшаяся полипептидная
цепь и удлинняется еще на одно звено.
Аминокислоты в растущей цепи соединены
в той последовательности, в которой
расположены шифрующие их кодоны в иРНК.
Для
того чтобы аминокислота включилась в
полипептидную цепь, она должна оторваться
от тРНК. Это становится возможным, когда
тРНК поступает на рибососму и антикодон
узнает свой кодон в иРНК. В рибосоме
имеется два участка для связывания двух
молекул тРНК. В один из этих участков,
называемый акцепторным
,
поступает тРНК с аминокислотой и
присоединяется к своему кодону (I).
Эта аминокислота присоединяет к себе
(акцептирует) растущую цепь белка (II)?
Между ними образуется пептидная связь.
тРНК, к которой теперь присоединяется
вместе с кодоном иРНК в донорный
участок рибосомы. В освободившийся
акцепторный участок приходит новая
тРНК, связанная с аминокислотой, которая
шифруется очередным кодоном (III).
Из донорного участка сюда вновь
переносится оторвавшаяся полипептидная
цепь и удлинняется еще на одно звено.
Аминокислоты в растущей цепи соединены
в той последовательности, в которой
расположены шифрующие их кодоны в иРНК.
Когда на рибосоме оказывается один из трех триплетов (УАА, УАГ, УГА ), являющиеся «знаками препинания» между генами, ни одна тРНК не может занять место в акцепторном участке. Дело в том, что не существует антикодонов, комплементарных последовательностям нуклеотидов «знаков препинания». Оторвавшейся цепи не к чему присоединиться в акцепторном участке, и она покидает рибосому. Синтез белка завершен.
У прокариот синтез белков начинается с того, что кодон АУГ , расположенный на первом месте в копии с каждого гена, занимае в рибосоме такую позицию, что с ним взаимодействует антикодон особой тРНК, оединенной с формилментионином . Эта измененная форма аминокислоты метионина сразу попадает в донорный участок и выполняет роль заглавной буквы во фразе – с нее в бактериальной клетке начинается синтез любой полипептидной цепи. Когда триплет АУГ стоит не на первом месте, а внутри копии с гена, он кодирует аминокислоту метионин. После завершения синтеза полипептидной цепи формилметионин отщепляется от нее и в готовом белке отсуствует.
 Для
увеличения производства белков иРНК
часто проходит одновременно не по одной,
а по нескольким рибосомам. Акую структуру,
объединенную одной молекулой иРНК,
называют полисомой
.
На каждой рибосоме вэтом похожем на
нитку бус конвейере синтезируются
одинаковые белки.
Для
увеличения производства белков иРНК
часто проходит одновременно не по одной,
а по нескольким рибосомам. Акую структуру,
объединенную одной молекулой иРНК,
называют полисомой
.
На каждой рибосоме вэтом похожем на
нитку бус конвейере синтезируются
одинаковые белки.
Аминокислоты бесперебойно поставляются к рибосомам с помощью тРНК. Отдав аминокислоту, тРНК покидает рибосому и с помощью кодазы соединяется. Высокая слаженность всех «служб комбината» по производсву белов позволяет в течении нескольких секунд синтезировать полипептидные цепи, состоящие из сотен аминокислот.
Свойства генетического кода. Благодаря процессу транскрипции в клетке осуществляется передача информации от ДНК к белку
ДНК → иРНК → белок
Генетическая информация, содержащаяся в ДНК и в иРНК, заключена в последовательности расположения нуклеотидов в молекулах.
 Каким
же образом происходит перевод информации
с «языка» нуклеотидов на «язык»
аминокислот? Такой перевод осуществляется
с помощью генетического кода. Код,
или шифр
,
- это система символов для перевода
одной формы информации в другую.
Генетический
код
–это
система записи информации о
последовательности расположения
аминокислот в белках с помощью
последовательности расположения
нуклеотидов в иРНК.
Каким
же образом происходит перевод информации
с «языка» нуклеотидов на «язык»
аминокислот? Такой перевод осуществляется
с помощью генетического кода. Код,
или шифр
,
- это система символов для перевода
одной формы информации в другую.
Генетический
код
–это
система записи информации о
последовательности расположения
аминокислот в белках с помощью
последовательности расположения
нуклеотидов в иРНК.
Какими же свойствами обладает генетический код?
Код триплетен . В состав РНК входят четыре нуклеотида: А, Г, Ц, У. Если бы мы пытались обозначить одну аминокислоту одним нуклеотидом, то 16 из 20 аминокислот остались бы не зашифрованы. Двух буквенный код позволил бы зашифровать 16 аминокислот. Природа создала трехбуквенный, или триплетный, код. Это означает, что каждая из 20 аминокислот зашифрована последовательностью из трех нуклеотидов, называемой триплетом или кодоном.
Код вырожден. Это означает, что каждая аминокислота шифруется более чем одним кодоном. Исключения: метеонин и триптофан, каждая из которых кодируется одним триплетом.
Код однозначен. Каждый кодон шифрует только одну аминокислоту.
Между генами имеется «знаки препинания». В печатном тексте в конце каждой фразы стоит точка. Несколько связанных по смыслу фраз составляют абзац. На языке генетической информации таким абзацем являетсяоперон и комплементарная ему иРНК. Каждый ген в опероне прокариот или отдельный ген эукариот кодирует одну полипептидную цепочку – фразу. Так как в ряде случаев по матрице иРНК последовательно создается несколько разных полипептидных цепей, они должны быть отделены друг от друга. Для этого в генетическом годе имеются три специальных триплета – УАА, УАГ, УГА, каждый из которых обозначает прекращение синтеза одной полипептидной цепи. Таким образом, эти триплеты выполняют функцию знаков препинания. Они находятся в конце каждого гена.
Внутри гена нет «знаков препинания».
Код универсален. Генетический код един для всех живущих на Земле существ. У бактерий и грибов, пшеницы и хлопка, рыб и червей, лягушек и человека одни и те же триплеты кодируют одни и те же аминокислоты.

Принципы репликации ДНК. Преемственность генетического материала в поколениях клеток и организмов обеспечивается процессом репликации – удвоения молекул ДНК. Этот сложный процесс осуществляется комплексом нескольких ферментов и не обладающих каталитической активностью белов, необходимых для придания полинуклеотидным цепям нужной конформации. В результате репликации образуются две идентичные двойные спирали ДНК. Эти так называемые дочерние молекулы ничем не отличаются друг от друга и от исходной материнской молекулы ДНК. Репликация происходит в клетке перед делением, поэтому каждая дочерняя клетка получает точно такие же молекулы ДНК, какие имела материнская клетка. Процесс репликации основан на ряде принципов:
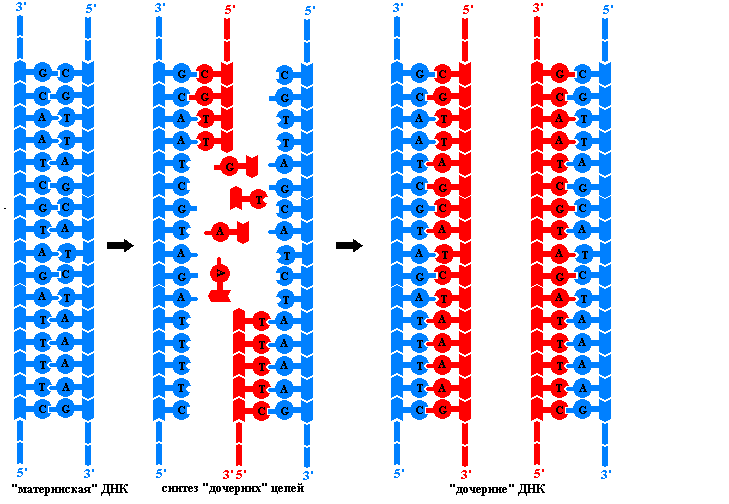
Только
в этом случае ДНК-полимеразы способна
двигаться по материнским нитям и
использовать их в качестве матриц для
безошибочного синтеза дочерних цепей.
Но полное раскручивание спиралей,
состоящих из многих миллионов пар
нуклеотидов, сопряжено со столь
значительным числом вращений и такими
энергетическими затратами, которые
невозможны в условиях клетки. Поэтому
репликация у эукариот начинается
одновременно в некоторых местах молекулы
ДНК. Участок между двумя точками, в
которых начинается синтез дочерних
цепей, называют репликоном
.
Он является единицей
репликации.


В каждой молекуле ДНК эукариотической клетки имеется много репликонов. В каждом репликоне можно видеть репликативную вилку – ту часть молекулы ДНК, которая под действием специальных ферментов уже расплелась. Каждая нить в вилке служит матрицей для синтеза комплементарной дочерней цепи. В ходе репликации вилка перемещается вдоль материнской молекулы, при этом расплетаются новые участки ДНК. Так как ДНК-полимеразы могут двигаться лишь в одном направлении вдоль матричных нитей, а нити ориентированы антипараллельно, то в каждой вилке одновременно ведут синтез два разных ферментативных комплекса. Причем в каждой вилке одна дочерняя (лидирующая) цепь растет непрерывно, а другая (отстающая) синтезируется отдельными фрагментами длинной в несколько нуклеотидов. Такие ферменты, названые в честь открывшего их японского ученого фрагментами Оказаки , сшиваются ДНК-лигазой, образуя непрерывную цепь. Механизм образования дочерних цепей ДНК фрагментами называют прерывистыми.
Потребность в затравке ДНК-полимераза не способна начинать синтез лидирующей цепи, ни синтез фрагментов Оказаки отстающей цепи. Она может лишь наращивать уже имеющуюся полинуклеотидную нить, последовательно присоединяя дезоксирибонуклеотиды к ее 3’-ОН концу. Откуда же берется начальный 5’-концевой участок растущей цепи ДНК? Его синтезирует на матрице ДНК особая РНК-полимераза, называемая праймазой (англ. Primer – затравка). Размер рибонуклеотидной затравки невелик (менее 20 нуклеотидов) в сравнении с размером цепи ДНК, образуемой ДНК-поимеразой. Выполнившая сво. Функци. РНК-затравка удаляется специальным ферментом, а образованная при эом брешь заделывается ДНК-полимеразой, использующей в качестве затравки 3’-ОН конец соседнего фрагмента Оказаки.
Проблема недорепликации концов линейных молекул ДНК. Удаление крайних РНК-праймеров, комплементрных 3’-концам обеих цепей линейной материнской молекулы ДНК, приводит к тому, что дочерние цепи оказываются короче 10-20 нуклеотидов. В этом и заключается проблема недорепликации концов линейных молекул.
Проблема недорепликации 3’-концов линейных молекул ДНК решается эукариотическими клетками с помощью специального фермента – теломеразы .
Теломераза является ДНК-полимеразой, достраивающей 3’-концылинейных молекул ДНК хромосом короткими повторяющимися последовательностями. Они, располагаясь друг за другом, образуют регулярную концевую структуру длинной до 10 тыс. нуклеотидов. Помимо белковой части, теломераза содержит РНК, выполняющую роль матрицы для наращивания ДНК повторами.
Схема удлинения концов молекул ДНК. Сначала происходит комплементарное связывание выступающего конца ДНК с матричным участком теломеразной РНК, затем теломераза наращивает ДНК, используя в качестве затравки ее 3’-ОН конец, а в качестве матрицы – РНК, входящую в состав фермента. Эта стадия называется элонгацией. После этого происходит транслокация, т.е. перемещение ДНК, удлиненной на один повтор, относительно фермента. Следом идет элонгация и очередная транслокация.
В результате образуются специализированные концевые структуры хромосом. Они состоят из многократно повторенных коротких последовательностей ДНК и специфических белков.
Рибонуклеиновая кислота представляет собой сополимер пуриновых и пиримидиновых рибонуклеотидов, соединенных друг с другом, как и в ДНК, -фосфодиэфирными мостиками (рис. 37.6). Хотя эти два вида нуклеиновых кислот имеют много общего, по ряду признаков они отличаются друг от друга.
1. У РНК углеводным остатком, к которому присоединены пуриновые или пиримидиновые основания и фосфатные группы, является рибоза, а не 2-дезоксирибоза (как у ДНК).
2. Пиримидиновые компоненты РНК отличаются от таковых у ДНК. В состав РНК, как и в состав ДНК, входят нуклеотиды аденина, гуанина и цитозина. В то же время РНК (за исключением некоторых специальных случаев, на которых мы остановимся ниже) не содержит тимина, его место в молекуле РНК занимает урацил.
3. РНК - одноцепочечная молекула (в отличие от ДНК, имеющей двухцепочечную структуру), однако при наличии в цепи РНК участков с комплементарной последовательностью (противоположной полярности) единичная цепь РНК способна сворачиваться с образованием так называемых «шпилек», структур, имеющих двухспиральные характеристики (рис. 37.7).

Рис. 37.6. Фрагмент молекулы рибонуклеиновой кислоты (РНК), в котором пуриновые и пиримидиновые основания- аденин (А), урацил (U), цитозин (С) и гуанин (-удерживаются фосфодиэфирным остовом, соединяющим рибозильные остатки, связанные N-гликозидной связью с соответствующими нуклеиновыми основаниями. Обратите внимание: цепь РНК обладает определенной направленностью, на которую указывают 5- и З-концевые фосфатные остатки.
4. Так как молекула РНК представляет собой одиночную цепь, комплементарную только одной из цепей ДНК, содержание в ней гуанина не обязательно равно содержанию цитозина, а содержание аденина не обязательно равно содержанию урацила.
5. РНК может быть гидролизована щелочью до 2, З-циклических диэфиров мононуклеотидов; в роли промежуточного продукта гидролиза выступает 2, У, 5-триэфир, который не образуется при щелочном гидролизе ДНК из-за отсутствия у последней 2-гидроксильных групп; щелочная лабильность РНК (сравнительно с ДНК) является полезным свойством как для диагностических, так и для аналитических целей.
Информация, содержащаяся в одноцепочечной РНК, реализуется в виде определенной последовательности пуриновых и пиримидиновых оснований (т. е. в первичной структуре) полимерной цепи. Эта последовательность комплементарна кодирующей цепи гена, с которой «считывается» РНК. Вследствие комплементарности молекула РНК способна специфически связываться (гибридизоваться) с кодирующей цепью, но не гибридизуется с некодирующей цепью ДНК. Последовательность РНК (за исключением замены Т на U) идентична последовательности некодирующей цепи гена (рис. 37.8).
Биологические функции РНК
Известно несколько видов РНК. Почти все они непосредственно вовлечены в процесс биосинтеза белка. Молекулы цитоплазматической РНК, выполняющие функции матриц белкового синтеза, называются матричными РНК (мРНК). Другой вид цитоплазматической РНК-рибосомная РНК (рРНК) - выполняет роль структурных компонентов рибосом (органелл, играющих важную роль в синтезе белка). Адапторные молекулы транспортных РНК (тРНК) участвуют в трансляции (переводе) информации мРНК в последовательность аминокислот в белках.
Значительная часть РНК-первичных транскриптов, образующихся в эукариотических клетках, включая и клетки млекопитающих, - подвергается деградации в ядре и не играет какой-либо структурной или информационной роли в цитоплазме. В культивируемых

Рис. 37.7. Вторичная структура молекулы РНК типа «петли со стеблем» («шпилька»), возникающая вследствие внутримолекулярного образования водородных связей между комплементарными парами нуклеиновых оснований.
клетках человека обнаружен класс малых ядерных РНК которые непосредственно не участвуют в синтезе белка, но могут оказывать влияние на процессинг РНК и общую «архитектуру» клетки. Размеры этих относительно небольших молекул варьируют, последние содержат от 90 до 300 нуклеотидов (табл. 37.3).
РНК является основным генетическим материалом у некоторых вирусов животных и растений. Некоторые РНК-содержащие вирусы никогда не проходят стадию обратной транскрипции РНК в ДНК. Однако для большинства известных вирусов животных, таких, как ретровирусы, характерна обратная транскрипция их РНК-генома, направляемая РНК-зависимой ДНК-полимеразой (обратной транскриптазой) с образованием двухспиральной ДНК-копии. Во многих случаях образующийся двухспиральный ДНК-транскрипт встраивается в геном и в дальнейшем обеспечивает экспрессию генов вируса, а также наработку новых копий вирусных РНК-геномов.
Структурная организация РНК
Во всех эукариотических и прокариотических организмах существуют три основных класса молекул РНК: информационная (матричная или мессенджер) РНК (мРНК), транспортная (тРНК) и рибосомная (рРНК). Представители этих классов отличаются друг от друга размерами, функциями и стабильностью.
Информационная (мРНК) - наиболее гетерогенный в отношении размеров и стабильности класс. Все представители этого класса служат переносчиками информации от гена к белок-синтезирующей системе клетки. Они выполняют роль матриц для синтезируемого полипептида, т. е. определяют аминокислотную последовательность белка (рис. 37.9).
Информационные РНК, особенно эукариотические, обладают некоторыми уникальными структурными особенностями. 5-Конец мРНК «кэпирован» 7-метилгуанозинтрифосфатом, присоединенным к 5-гидроксилу соседнего 2-0-метилрибонуклеозида через остаток трифосфата (рис. 37.10). Молекулы мРНК часто содержат внутренние остатки 6-метиладенина и 2-0-метилированные рибонуклеотиды. Хотя смысл «кэпирования» до конца еще не выяснен, можно предположить, что образующаяся структура 5-конца мРНК используется для специфического узнавания в системе трансляции. Синтез белка начинается на 5"-(кэпированном) конце мРНК. Другой конец большинства молекул мРНК (З-конец) содержит полиаденилатную цепочку из 20-250 нуклеотидов. Специфические функции этого окончательно не установлены. Можно предполагать, что данная структура отвечает за поддержание внутриклеточной стабильности мРНК. Некоторые мРНК, включая гистоновые не содержат poly (А). Наличие poly (А) в структуре мРНК используется для отделения от других видов РНК посредством фракционирования тотальной РНК на колонках с oligo (Т), иммобилизованным на твердом носителе типа целлюлозы. Связывание мРНК с колонкой происходит за счет комплементарных взаимодействий poly (А)-«хвоста» с иммобилизованным oligo (Т).
Рис. 37.8. Последовательность гена и его РНК-транскрипта. Показаны кодирующая и некодирующая цепи, и отмечена их полярность. РНК-транскрипт, имеющий полярность комплементарен кодирующей цепи (с полярностью 3 - 5) и идентичен по последовательности (за исключением замен Т на U) и полярности некодирующей цепи ДНК.

Рис. 37.9. Экспрессия генетической информации ДНК в форме мРНК-транскрипта и последующая трансляция при участии рибосом с образованием специфической молекулы белка.
(см. скан)
Рис. 37.10. Структура «кэпа», находящегося на 5-конце большинства эукариотических матричных РНК 7-метилгуанозинтрифосфат присоединяется к 5-концу мРНК. на котором обычно находится 2-О-метилпуриновый нуклеотид.
В клетках млекопитающих, включая клетки человека, зрелые молекулы мРНК, находящиеся в цитоплазме, не являются полной копией транскрибируемого участка гена. Образующийся в результате транскрипции полирибонуклеотид представляет собой предшественник цитоплазматической мРНК, перед выходом из ядра он подвергается специфическому процессингу. Непроцессированные продукты транскрипции, обнаруживаемые в ядрах клеток млекопитающих, образуют четвертый класс молекул РНК. Такие ядерные РНК очень гетерогенны и достигают значительных размеров. Молекулы гетерогенных ядерных РНК могут иметь молекулярную массу более , в то время как молекулярная масса мРНК обычно не превышает 2106. подвергаются процессингу в ядре, и образующиеся зрелые мРНК поступают в цитоплазму, где служат матрицей для биосинтеза белка.
Молекулы транспортных РНК (тРНК) обычно содержат около 75 нуклеотидов. Молекулярная масса таких молекул составляет . тРНК также формируются в результате специфического процессинга соответствующих молекул-предшественников (см. гл. 39). Транспортные тРНК выполняют функцию посредников в ходе трансляции мРНК. В любой клетке присутствуют не менее 20 видов молекул тРНК. Каждый вид (иногда несколько видов) тРНК соответствует одной из 20 аминокислот, необходимых для синтеза белка. Хотя каждая специфическая тРНК отличается от других нуклеотидной последовательностью, все они имеют и общие черты. Благодаря нескольим внутрицепочечным комплементарным участкам, все тРНК обладают вторичной структурой, получившей название «клеверный лист» (рис. 37.11).
Молекулы всех видов тРНК имеют четыре основных плеча. Акцепторное плечо состоит из «стебля» спаренных нуклеотидов и заканчивается последовательностью ССА Именно через У-гидроксильную группу аденозильного остатка происходит связывание с карбоксильной группой аминокислоты. Остальные плечи тоже состоят из «стеблей», образованных комплементарными парами оснований, и петель из неспаренных оснований (рис. 37.7). Антикодоновое плечо узнает нуклеотидный триплет или кодон (см. гл. 40) в мРНК. D-плечо названо так из-за наличия в нем дигидроуридина, -плечо названо по последовательности Т-псевдоуридин-С. Дополнительное плечо представляет собой наиболее вариабельную структуру и служит основой классификации тРНК. тРНК класса 1 (75% от общего их числа) обладают дополнительным плечом длиной 3-5 пар оснований. Дополнительное плечо у тРНК-молекул класса 2 состоит из 13-21 пар оснований и часто включает неспаренную петлю.

Рис. 37.11. Структура молекулы аминоацил-тРНК, к 3-ССА-концу которой присоединена аминокислота . Указаны внутримолекулярные водородные связи и расположение антикодонового, ТТС- и дигидроурацилового плеч. (From J. D. Watson. Molecular biology of the Gene 3rd, ed.. Copyright 1976, 1970, 1965 by W. A. Benjamin, Inc., Menlo Park Calif.)
Вторичная структура, определяемая системой комплементарных взаимодействий нуклеотидных оснований соответствующих плеч, характерна для всех видов Акцепторное плечо содержит семь пар оснований, -плечо - пять пар оснований, плечо D - три (или четыре) пары оснований.
Молекулы тРНК весьма стабильны у прокариот и несколько менее стабильны у эукариот. Обратная ситуация характерна для мРНК, которая довольно нестабильна у прокариот, а у эукариотических организмов обладает значительной стабильностью.
Рибосомная РНК. Рибосома - это цитоплазматическая нуклеопротеиновая структура, предназначенная для синтеза белка по мРНК-матрице. Рибосома обеспечивает специфический контакт в результате которого и происходит трансляция нуклеотидной последовательности, считанной с определенного гена, в аминокислотную последовательность соответствующего белка.
В табл. 37.2 представлены компоненты рибосом млекопитающих, имеющих молекулярную массу 4,210 6 и скорость седиментации (единиц Сведберга). Рибосомы млекопитающих состоят из двух нуклеопротеиновых субъединиц - большой с
Таблица 37.2. Компоненты рибосом млекопитающих

молекулярной массой (60S), и малой, имеющей молекулярную массу (40S). 608-субъединица содержит 58-рибосомную РНК (рРНК), 5,8S-pPHK и 28S-pPHK, а также более 50 различных полипептидов. Малая, 408-субъединица включает единственную 18S-pPHK и около 30 полипептидных цепей. Все рибосомные РНК, за исключением 5S-PHK, имеют общего предшественника-45S-PHK, локализованную в ядрышке (см. гл. 40). У молекулы 5S-PHK предшественник собственный. В ядрышке происходит упаковка высокометилированных рибосомных РНК с рибосомными белками. В цитоплазме рибосомы достаточно устойчивы и способны осуществлять большое число циклов трансляции.
Небольшие стабильные РНК. В эукариотических клетках обнаружено большое число дискретных, высококонсервативных, небольших и стабильных молекул РНК. Большинство РНК этого типа обнаруживаются в составе рибонуклеопротеинов и локализованы в ядре, цитоплазме или одновременно в обоих компартментах. Размеры этих молекул варьируют от 90 до 300 нуклеотидов, содержание их - 100000-1000000 копий на клетку.
Малые ядерные нуклеопротеиновые частицы (часто называемые snurps - от англ. small nuclear ribonucleic particles), вероятно, играют существенную роль в регуляции экспрессии генов. Нуклеопротеиновые частицы типа U7, по-видимому, участвуют в формировании З-концов гистоновых мРНК. Частицы , вероятно, необходимы для полиаденилирования, a - для удаления интронов и процессинга мРНК (см. гл. 39). Табл. 37.3. суммирует некоторые характеристики небольших стабильных РНК.
Таблица 37.3. Некоторые виды небольших стабильных РНК, обнаруженные в клетках млекопитающих

ЛИТЕРАТУРА
Darnell J. et al. Molecular Cell Biology, Scientific American Books, 1986.
Hunt T. DNA Makes RNA Makes Protein, Elsevier, 1983. Lewin B. Genes, 2nd ed., Wiley, 1985.
Rich A. et al. The chemistry and biology of left-handed Z-DNA, Annu. Rev. Biochem., 1984, 53, 847.
Turner P. Controlling roles for snurps, Nature, 1985, 316, 105. Watson J. D. The Double Helix, Atheneum, 1968.
Watson J. D., Crick F.H.C. Molecular structure of nucleic acids. Nature, 1953, 171, 737.
Zieve G. W. Two groups of small stable RNAs, Cell, 1981, 25, 296.
Справа крупнейшая спираль ДНК человека, выстроенная из людей на пляже в Варне (Болгария), вошедшая в книгу рекордов Гиннесса 23 апреля 2016 года
Дезоксирибонуклеиновая кислота. Общие сведения
ДНК (дезоксирибонуклеиновая кислота) - своеобразный чертеж жизни, сложный код, в котором заключены данные о наследственной информации. Эта сложная макромолекула способна хранить и передавать наследственную генетическую информацию из поколения в поколение. ДНК определяет такие свойства любого живого организма как наследственность и изменчивость. Закодированная в ней информация задает всю программу развития любого живого организма. Генетически заложенные факторы предопределяют весь ход жизни как человека, так и любого др. организхма. Искусственное или естественное воздействие внешней среды способны лишь в незначительной степени повлиять на общую выраженность отдельных генетических признаков или сказаться на развитии запрограммированных процессов.
Дезоксирибонуклеи́новая кислота (ДНК) — макромолекула (одна из трёх основных, две другие — РНК и белки), обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. ДНК содержит информацию о структуре различных видов РНК и белков.
В клетках эукариот (животных, растений и грибов) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами.
С химической точки зрения ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков — нуклеотидов. Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы. Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы (С ) и фосфатной (Ф ) группы (фосфодиэфирные связи).

Рис. 2. Нуклертид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы
В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей, ориентированных азотистыми основаниями друг к другу. Эта двухцепочечная молекула закручена по винтовой линии.
В ДНК встречается четыре вида азотистых оснований (аденин, гуанин, тимин и цитозин). Азотистые основания одной из цепей соединены с азотистыми основаниями другой цепи водородными связями согласно принципу комплементарности: аденин соединяется только с тимином (А-Т ), гуанин — только с цитозином (Г-Ц ). Именно эти пары и составляют «перекладины» винтовой "лестницы" ДНК (см.: рис. 2, 3 и 4).

Рис. 2. Азотистые основания
Последовательность нуклеотидов позволяет «кодировать» информацию о различных типах РНК, наиболее важными из которых являются информационные, или матричные (мРНК), рибосомальные (рРНК) и транспортные (тРНК). Все эти типы РНК синтезируются на матрице ДНК за счёт копирования последовательности ДНК в последовательность РНК, синтезируемой в процессе транскрипции, и принимают участие в биосинтезе белков (процессе трансляции). Помимо кодирующих последовательностей, ДНК клеток содержит последовательности, выполняющие регуляторные и структурные функции.

Рис. 3. Репликация ДНК
Расположение базовых комбинаций химических соединений ДНК и количественные соотношения между этими комбинациями обеспечивают кодирование наследственной информации.
Образование новой ДНК (репликация)
- Процесс репликации: раскручивание двойной спирали ДНК — синтез комплементарных цепей ДНК-полимеразой — образование двух молекул ДНК из одной.
- Двойная спираль «расстегивается» на две ветви, когда ферменты разрушают связь между базовыми парами химических соединений.
- Каждая ветвь является элементом новой ДНК. Новые базовые пары соединяются в той же последовательности, что и в родительской ветви.
По завершении дупликации образуются две самостоятельные спирали, созданные из химических соединений родительской ДНК и имеющие с ней одинаковый генетический код. Таким путем ДНК способна перерывать информацию от клетки к клетке.
Более подробная информация:
СТРОЕНИЕ НУКЛЕИНОВЫХ КИСЛОТ

Рис. 4 . Азотистые основания: аденин, гуанин, цитозин, тимин
Дезоксирибонуклеиновая кислота (ДНК) относится к нуклеиновым кислотам. Нуклеиновые кислоты - это класс нерегулярных биополимеров, мономерами которых являются нуклеотиды.
НУКЛЕОТИДЫ состоят из азотистого основания , соединенного с пятиуглеродным углеводом (пентозой) - дезоксирибозой (в случае ДНК) или рибозой (в случае РНК), который соединяется с остатком фосфорной кислоты (H 2 PO 3 -).
Азотистые основания бывают двух типов: пиримидиновые основания - урацил (только в РНК), цитозин и тимин, пуриновые основания - аденин и гуанин.

Рис. 5. Структура нуклеотидов (слева), расположение нуклеотида в ДНК (снизу) и типы азотистых оснований (справа): пиримидиновые и пуриновые

Атомы углерода в молекуле пентозы нумеруются числами от 1 до 5. Фосфат соединяется с третьим и пятым атомами углерода. Так нуклеинотиды соединяются в цепь нуклеиновой кислоты. Таким образом, мы можем выделить 3’ и 5’-концы цепи ДНК:

Рис. 6. Выделение 3’ и 5’-концов цепи ДНК
Две цепи ДНК образуют двойную спираль . Эти цепи в спирали сориентированы в противоположных направлениях. В разных цепях ДНК азотистые основания соединены между собой с помощью водородных связей . Аденин всегда соединяется с тимином, а цитозин - с гуанином. Это называется правилом комплементарности .
Правило комплементарности:
| A-T G-C |
Например, если нам дана цепь ДНК, имеющая последовательность
3’- ATGTCCTAGCTGCTCG - 5’,
то вторая ей цепь будет комплементарна и направлена в противоположном направлении - от 5’-конца к 3’-концу:
5’- TACAGGATCGACGAGC- 3’.

Рис. 7. Направленность цепей молекулы ДНК и соединение азотистых оснований с помощью водородных связей
РЕПЛИКАЦИЯ ДНК
Репликация ДНК - это процесс удвоения молекулы ДНК путем матричного синтеза. В большинстве случаев естественной репликации ДНК праймером для синтеза ДНК является короткий фрагмент (создаваемый заново). Такой рибонуклеотидный праймер создается ферментом праймазой (ДНК-праймаза у прокариот, ДНК-полимераза у эукариот), и впоследствии заменяется дезоксирибонуклеотидами полимеразой, выполняющей в норме функции репарации (исправления химических повреждений и разрывов в молекле ДНК).
Репликация происходит по полуконсервативному механизму. Это значит, что двойная спираль ДНК расплетается и на каждой из ее цепей по принципу комплементарности достраивается новая цепь. Дочерняя молекула ДНК, таким образом, содержит в себе одну цепь от материнской молекулы и одну вновь синтезированную. Репликация происходит в направлении от 3’ к 5’ концу материнской цепи.

Рис. 8. Репликация (удвоение) молекулы ДНК
ДНК-синтез - это не такой сложный процесс, как может показаться на первый взгляд. Если подумать, то для начала нужно разобраться, что же такое синтез. Это процесс объединения чего-либо в одно целое. Образование новой молекулы ДНК проходит в несколько этапов:
1) ДНК-топоизомераза, располагаясь перед вилкой репликации, разрезает ДНК для того, чтобы облегчить ее расплетание и раскручивание.
2) ДНК-хеликаза вслед за топоизомеразой влияет на процесс «расплетения» спирали ДНК.
3) ДНК-связывающие белки осуществляют связывание нитей ДНК, а также проводят их стабилизацию, не допуская их прилипания друг к другу.
4) ДНК-полимераза δ
(дельта), согласовано со скоростью движения репликативной вилки, осуществляет синтез
ведущей
цепи
дочерней ДНК в направлении 5"→3" на матрице
материнскойнити ДНК по направлению от ее 3"-конца к 5"-концу (скорость до 100 пар нуклеотидов в секунду). Этим события на данной материнской
нити ДНК ограничиваются.

Рис. 9. Схематическое изображение процесса репликации ДНК: (1) Отстающая цепь (запаздывающая нить), (2) Ведущая цепь (лидирующая нить), (3) ДНК-полимераза α (Polα ), (4) ДНК-лигаза, (5) РНК-праймер, (6) Праймаза, (7) Фрагмент Оказаки, (8) ДНК-полимераза δ (Polδ ), (9) Хеликаза, (10) Однонитевые ДНК-связывающие белки, (11) Топоизомераза.
Далее описан синтез отстающей цепи дочерней ДНК (см. Схему репликативной вилки и функции ферментов репликации)
Нагляднее о репликации ДНК см.
5) Непосредственно сразу после расплетания и стабилизации другой нити материнской молекулы к ней присоединяется ДНК-полимераза α (альфа) и в направлении 5"→3" синтезирует праймер (РНК-затравку) - последовательность РНК на матрице ДНК длиной от 10 до 200 нуклеотидов. После этого фермент удаляется с нити ДНК.
Вместо
ДНК-полимеразы
α
к 3"-концу праймера присоединяется
ДНК-полимераза
ε
.
6)
ДНК-полимераза
ε
(эпсилон)
как бы продолжает удлинять праймер, но в качестве субстрата встраивает
дезоксирибонуклеотиды
(в количестве 150-200 нуклеотидов). В результате образуется цельная нить из двух частей -
РНК
(т.е. праймер) и
ДНК
.
ДНК-полимераза
ε
работает до тех пор, пока не встретит праймер предыдущего
фрагмента Оказаки
(синтезированный чуть ранее). После этого данный фермент удаляется с цепи.
7) ДНК-полимераза β (бета) встает вместо ДНК-полимеразы ε , движется в том же направлении (5"→3") и удаляет рибонуклеотиды праймера, одновременно встраивая дезоксирибонуклеотиды на их место. Фермент работает до полного удаления праймера, т.е. пока на его пути не встанет дезоксирибонуклеотид (еще более ранее синтезированный ДНК-полимеразой ε ). Связать результат свой работы и впереди стоящую ДНК фермент не в состоянии, поэтому он сходит с цепи.
В результате на матрице материнской нити "лежит" фрагмент дочерней ДНК. Он называется
фрагмент Оказаки
.
8) ДНК-лигаза производит сшивку двух соседних фрагментов Оказаки , т.е. 5"-конца отрезка, синтезированного ДНК-полимеразой ε , и 3"-конца цепи, встроенного ДНК-полимеразой β .
СТРОЕНИЕ РНК
Рибонуклеиновая кислота (РНК) — одна из трёх основных макромолекул (две другие — ДНК и белки), которые содержатся в клетках всех живых организмов.
Так же, как ДНК, РНК состоит из длинной цепи, в которой каждое звено называется нуклеотидом . Каждый нуклеотид состоит из азотистого основания, сахара рибозы и фосфатной группы. Однако в отличие от ДНК, РНК обычно имеет не две цепи, а одну. Пентоза в РНК представлена рибозой, а не дезоксирибозой (у рибозы присутствует дополнительная гидроксильная группа на втором атоме углевода). Наконец, ДНК отличается от РНК по составу азотистых оснований: вместо тимина (Т ) в РНК представлен урацил (U ) , который также комплементарен аденину.
Последовательность нуклеотидов позволяет РНК кодировать генетическую информацию. Все клеточные организмы используют РНК (мРНК) для программирования синтеза белков.
Клеточные РНК образуются в ходе процесса, называемого транскрипцией , то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами - РНК-полимеразами .
Затем матричные РНК (мРНК) принимают участие в процессе, называемом трансляцией, т.е. синтеза белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.

Рис. 10. Отличие ДНК от РНК по азотистому основанию: вместо тимина (Т) в РНК представлен урацил (U), который также комплементарен аденину.
ТРАНСКРИПЦИЯ
Это процесс синтеза РНК на матрице ДНК. ДНК раскручивается на одном из участков. На одной из цепей содержится информация, которую необходимо скопировать на молекулу РНК - эта цепь называется кодирующей. Вторая цепь ДНК, комплементарная кодирующей, называется матричной. В процессе транскрипции на матричной цепи в направлении 3’ - 5’ (по цепи ДНК) синтезируется комплементарная ей цепь РНК. Таким образом, создается РНК-копия кодирующей цепи.
![]()
Рис. 11. Схематическое изображение транскрипции
Например, если нам дана последовательность кодирующей цепи
3’- ATGTCCTAGCTGCTCG - 5’,
то, по правилу комплементарности, матричная цепь будет нести последовательность
5’- TACAGGATCGACGAGC- 3’,
а синтезируемая с нее РНК - последовательность
ТРАНСЛЯЦИЯ
Рассмотрим механизм синтеза белка
на матрице РНК, а также генетический код и его свойства. Также для наглядности по ниже приведенной ссылке рекомендуем посмотреть небольшое видео о процессах транскрипции и трансляции, происходящих в живой клетке:

Рис. 12. Процесс синтеза белка: ДНК кодирует РНК, РНК кодирует белок
ГЕНЕТИЧЕСКИЙ КОД
Генетический код - способ кодирования аминокислотной последовательности белков с помощью последовательности нуклеотидов. Каждая аминокислота кодируется последовательностью из трех нуклеотидов - кодоном или триплетом.
Генетический код, общий для большинства про- и эукариот. В таблице приведены все 64 кодона и указаны соответствующие аминокислоты. Порядок оснований — от 5" к 3" концу мРНК.
Таблица 1. Стандартный генетический код
|
1-е
ние |
2-е основание |
3-е
ние |
|||||||
|
U |
C |
A |
G |
||||||
|
U |
U U U |
(Phe/F) |
U C U |
(Ser/S) |
U A U |
(Tyr/Y) |
U G U |
(Cys/C) |
U |
|
U U C |
U C C |
U A C |
U G C |
C |
|||||
|
U U A |
(Leu/L) |
U C A |
U A A |
Стоп-кодон** |
U G A |
Стоп-кодон** |
A |
||
|
U U G |
U C G |
U A G |
Стоп-кодон** |
U G G |
(Trp/W) |
G |
|||
|
C |
C U U |
C C U |
(Pro/P) |
C A U |
(His/H) |
C G U |
(Arg/R) |
U |
|
|
C U C |
C C C |
C A C |
C G C |
C |
|||||
|
C U A |
C C A |
C A A |
(Gln/Q) |
C GA |
A |
||||
|
C U G |
C C G |
C A G |
C G G |
G |
|||||
|
A |
A U U |
(Ile/I) |
A C U |
(Thr/T) |
A A U |
(Asn/N) |
A G U |
(Ser/S) |
U |
|
A U C |
A C C |
A A C |
A G C |
C |
|||||
|
A U A |
A C A |
A A A |
(Lys/K) |
A G A |
A |
||||
|
A U G |
(Met/M) |
A C G |
A A G |
A G G |
G |
||||
|
G |
G U U |
(Val/V) |
G C U |
(Ala/A) |
G A U |
(Asp/D) |
G G U |
(Gly/G) |
U |
|
G U C |
G C C |
G A C |
G G C |
C |
|||||
|
G U A |
G C A |
G A A |
(Glu/E) |
G G A |
A |
||||
|
G U G |
G C G |
G A G |
G G G |
G |
|||||
Среди триплетов есть 4 специальных последовательности, выполняющих функции «знаков препинания»:
- *Триплет AUG , также кодирующий метионин, называется старт-кодоном . С этого кодона начинается синтез молекулы белка. Таким образом, во время синтеза белка, первой аминокислотой в последовательности всегда будет метионин.
- **Триплеты UAA , UAG и UGA называются стоп-кодонами и не кодируют ни одной аминокислоты. На этих последовательностях синтез белка прекращается.
Свойства генетического кода
1. Триплетность . Каждая аминокислота кодируется последовательностью из трех нуклеотидов - триплетом или кодоном.
2. Непрерывность . Между триплетами нет никаких дополнительных нуклеотидов, информация считывается непрерывно.
3. Неперекрываемость . Один нуклеотид не может входить одновременно в два триплета.
4. Однозначность . Один кодон может кодировать только одну аминокислоту.

5. Вырожденность . Одна аминокислота может кодироваться несколькими разными кодонами.

6. Универсальность . Генетический код одинаков для всех живых организмов.
Пример. Нам дана последовательность кодирующей цепи:
3’- CCGATTGCACGTCGATCGTATA - 5’.
Матричная цепь будет иметь последовательность:
5’- GGCTAACGTGCAGCTAGCATAT - 3’.
Теперь «синтезируем» с этой цепи информационную РНК:
3’- CCGAUUGCACGUCGAUCGUAUA - 5’.
Синтез белка идет в направлении 5’ → 3’, следовательно, нам нужно перевернуть последовательность, чтобы «прочитать» генетический код:
5’- AUAUGCUAGCUGCACGUUAGCC - 3’.
Теперь найдем старт-кодон AUG:
5’- AUAUG CUAGCUGCACGUUAGCC - 3’.
Разделим последовательность на триплеты:
звучит следующим образом: информация с ДНК передается на РНК (транскрипция), с РНК - на белок (трансляция). ДНК также может удваиваться путем репликации, и также возможен процесс обратной транскрипции, когда по матрице РНК синтезируется ДНК, но такой процесс в основном характерен для вирусов.

Рис. 13. Центральная догма молекулярной биологии
ГЕНОМ: ГЕНЫ и ХРОМОСОМЫ
(общие понятия)
Геном - совокупность всех генов организма; его полный хромосомный набор.
Термин "геном" был предложен Г. Винклером в 1920 г. для описания совокупности генов, заключенных в гаплоидном наборе хромосом организмов одного биологического вида. Первоначальный смысл этого термина указывал на то, что понятие генома в отличие от генотипа является генетической характеристикой вида в целом, а не отдельной особи. С развитием молекулярной генетики значение данного термина изменилось. Известно, что ДНК, которая является носителем генетической информации у большинства организмов и, следовательно, составляет основу генома, включает в себя не только гены в современном смысле этого слова. Большая часть ДНК эукариотических клеток представлена некодирующими ("избыточными") последовательностями нуклеотидов, которые не заключают в себе информации о белках и нуклеиновых кислотах. Таким образом, основную часть генома любого организма составляет вся ДНК его гаплоидного набора хромосом.
Гены — это участки молекул ДНК, кодирующие полипептиды и молекулы РНК
За последнее столетие наше представление о генах существенно изменилось. Ранее геном называли участок хромосомы, кодирующий или определяющий один признак или фенотипическое (видимое) свойство, например цвет глаз.
В 1940 г. Джордж Бидл и Эдвард Тейтем предложили молекулярное определение гена. Ученые обрабатывали споры гриба Neurospora crassa рентгеновским излучением и другими агентами, вызывающими изменения в последовательности ДНК (мутации ), и обнаружили мутантные штаммы гриба, утратившие некоторые специфические ферменты, что в некоторых случаях приводило к нарушению целого метаболического пути. Бидл и Тейтем пришли к выводу, что ген — это участок генетического материала, который определяет или кодирует один фермент. Так появилась гипотеза «один ген — один фермент» . Позднее эта концепция была расширена до определения «один ген — один полипептид» , поскольку многие гены кодируют белки, не являющиеся ферментами, а полипептид может оказаться субъединицей сложного белкового комплекса.
На рис. 14 показана схема того, как триплеты нуклеотидов в ДНК определяют полипептид - аминокислотную последовательность белка при посредничестве мРНК. Одна из цепей ДНК играет роль матрицы для синтеза мРНК, нуклеотидные триплеты (кодоны) которой комплементарны триплетам ДНК. У некоторых бактерий и многих эукариот кодирующие последовательности прерываются некодирующими участками(так называемыми интронами ).
Современное биохимическое определение гена еще более конкретно. Генами называются все участки ДНК, кодирующие первичную последовательность конечных продуктов, к которым относятся полипептиды или РНК, обладающие структурной или каталитической функцией.
Наряду с генами ДНК содержит и другие последовательности, выполняющие исключительно регуляторную функцию. Регуляторные последовательности могут обозначать начало или конец генов, влиять на транскрипцию или указывать место инициации репликации или рекомбинации. Некоторые гены могут экспрессироваться разными путями, при этом один и тот же участок ДНК служит матрицей для образования разных продуктов.
Мы можем приблизительно рассчитать минимальный размер гена , кодирующего средний белок. Каждая аминокислота в полипептидной цепи кодируется последовательностью из трех нуклеотидов; последовательности этих триплетов (кодонов) соответствуют цепочке аминокислот в полипептиде, который кодируется данным геном. Полипептидная цепь из 350 аминокислотных остатков (цепь средней длины) соответствует последовательности из 1050 п.н. (пар нуклеотидов ). Однако многие гены эукариот и некоторые гены прокариот прерываются сегментами ДНК, не несущими информации о белке, и поэтому оказываются значительно длиннее, чем показывает простой расчет.
Сколько генов в одной хромосоме?
 Рис. 15. Вид хромосом в прокаритической (слева) и эукариотической клеках. Гистоны (Histones) — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация.
Рис. 15. Вид хромосом в прокаритической (слева) и эукариотической клеках. Гистоны (Histones) — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация.
Как известно, бактериальные клетки имеют хромосому в виде нити ДНК, уложенной в компактную структуру - нуклеоид. Хромосома прокариота Escherichia coli , чей геном полностью расшифрован, представляет собой кольцевую молекулу ДНК (на самом деле, это не правильный круг, а скорее петля без начала и конца), состоящую из 4 639 675 п.н. В этой последовательности содержится примерно 4300 генов белков и еще 157 генов стабильных молекул РНК. В геноме человека примерно 3,1 млрд пар нуклеотидов, соответствующих почти 29 000 генам, расположенным на 24 разных хромосомах.
Прокариоты (Бактерии).
 Бактерия E. coli
имеет одну двухцепочечную кольцевую молекулу ДНК. Она состоит из 4 639 675 п.н. и достигает в длину примерно 1,7 мм, что превышает длину самой клетки E. coli
приблизительно в 850 раз. Помимо крупной кольцевой хромосомы в составе нуклеоида многие бактерии содержат одну или несколько маленьких кольцевых молекул ДНК, свободно располагающихся в цитозоле. Такие внехромосомные элементы называют плазмидами
(рис. 16).
Бактерия E. coli
имеет одну двухцепочечную кольцевую молекулу ДНК. Она состоит из 4 639 675 п.н. и достигает в длину примерно 1,7 мм, что превышает длину самой клетки E. coli
приблизительно в 850 раз. Помимо крупной кольцевой хромосомы в составе нуклеоида многие бактерии содержат одну или несколько маленьких кольцевых молекул ДНК, свободно располагающихся в цитозоле. Такие внехромосомные элементы называют плазмидами
(рис. 16).
Большинство плазмид состоит всего из нескольких тысяч пар нуклеотидов, некоторые содержат более 10000 п. н. Они несут генетическую информацию и реплицируются с образованием дочерних плазмид, которые попадают в дочерние клетки в процессе деления родительской клетки. Плазмиды обнаружены не только в бактериях, но также в дрожжах и других грибах. Во многих случаях плазмиды не дают никаких преимуществ клеткам-хозяевам, и их единственная задача — независимое воспроизведение. Однако некоторые плазмиды несут полезные для хозяина гены. Например, содержащиеся в плазмидах гены могут придавать клеткам бактерий устойчивость к антибактериальным агентам. Плазмиды, несущие ген β-лактамазы, обеспечивают устойчивость к β-лактамным антибиотикам, таким как пенициллин и амоксициллин. Плазмиды могут переходить от клеток, устойчивых к антибиотикам, к другим клеткам того же или другого вида бактерий, в результате чего эти клетки также становятся резистентными. Интенсивное применение антибиотиков является мощным селективным фактором, способствующим распространению плазмид, кодирующих устойчивость к антибиотикам (а также транспозонов, которые кодируют аналогичные гены) среди болезнетворных бактерий, и приводит к появлению бактериальных штаммов с устойчивостью к нескольким антибиотикам. Врачи начинают понимать опасность широкого использования антибиотиков и назначают их только в случае острой необходимости. По аналогичным причинам ограничивается широкое использование антибиотиков для лечения сельскохозяйственных животных.
См. также: Равин Н.В., Шестаков С.В. Геном прокариот // Вавиловский журнал генетики и селекции, 2013. Т. 17. № 4/2. С. 972-984.
Эукариоты.
Таблица 2. ДНК, гены и хромосомы некоторых организмов
|
Общая ДНК, п.н. |
Число хромосом* |
Примерное число генов |
|
|
Escherichia coli (бактерия) |
4 639 675 |
4 435 |
|
|
Saccharomyces cerevisiae (дрожжи) |
12 080 000 |
16** |
5 860 |
|
Caenorhabditis elegans (нематода) |
90 269 800 |
12*** |
23 000 |
|
Arabidopsis thaliana (растение) |
119 186 200 |
33 000 |
|
|
Drosophila melanogaster (плодовая мушка) |
120 367 260 |
20 000 |
|
|
Oryza sativa (рис) |
480 000 000 |
57 000 |
|
|
Mus musculus (мышь) |
2 634 266 500 |
27 000 |
|
|
Homo sapiens (человек) |
3 070 128 600 |
29 000 |
Примечание. Информация постоянно обновляется; для получения более свежей информации обратитесь к сайтам, посвященным отдельным геномным проектам
* Для всех эукариот, кроме дрожжей, приводится диплоидный набор хромосом. Диплоидный
набор хромосом (от греч. diploos- двойной и eidos- вид) - двойной набор хромосом (2n), каждая из которых имеет себе гомологичную.
**Гаплоидный набор. Дикие штаммы дрожжей обычно имеют восемь (октаплоидный) или больше наборов таких хромосом.
***Для самок с двумя Х хромосомами. У самцов есть Х хромосома, но нет Y, т. е. всего 11 хромосом.
В клетке дрожжей, одних из самых маленьких эукариот, в 2,6 раза больше ДНК, чем в клетке E. coli (табл. 2). Клетки плодовой мушки Drosophila , классического объекта генетических исследований, содержат в 35 раз больше ДНК, а клетки человека — примерно в 700 раз больше ДНК, чем клетки E. coli. Многие растения и амфибии содержат еще больше ДНК. Генетический материал клеток эукариот организован в виде хромосом. Диплоидный набор хромосом (2n ) зависит от вида организма (табл. 2).
Например, в соматической клетке человека 46 хромосом (рис. 17 ). Каждая хромосома эукариотической клетки, как показано на рис. 17, а , содержит одну очень крупную двухспиральную молекулу ДНК. Двадцать четыре хромосомы человека (22 парные хромосомы и две половые хромосомы X и Y) различаются по длине более чем в 25 раз. Каждая хромосома эукариот содержит определенный набор генов.

Рис. 17. Хромосомы эукариот. а — пара связанных и конденсированных сестринских хроматид из хромосомы человека. В такой форме эукариотические хромосомы пребывают после репликации и в метафазе в процессе митоза. б — полный набор хромосом из лейкоцита одного из авторов книги. В каждой нормальной соматической клетке человека содержится 46 хромосом.
Если соединить между собой молекулы ДНК человеческого генома (22 хромосомы и хромосомы X и Y или Х и Х), получится последовательность длиной около одного метра. Прим.: У всех млекопитающих и других организмов с гетерогаметным мужским полом, у самок две X-хромосомы (XX), а у самцов — одна X-хромосома и одна Y-хромосома (XY).
Большинство клеток человека , поэтому общая длина ДНК таких клеток около 2м. У взрослого человека примерно 10 14 клеток, таким образом, общая длина всех молекул ДНК составляет 2・10 11 км. Для сравнения, окружность Земли — 4・10 4 км, а расстояние от Земли до Солнца — 1,5・10 8 км. Вот как удивительно компактно упакована ДНК в наших клетках!
В клетках эукариот есть и другие органеллы, содержащие ДНК, — это митохондрии и хлоропласты. Выдвигалось множество гипотез относительно происхождения ДНК митохондрий и хлоропластов. Общепризнанная сегодня точка зрения заключается в том, что они представляют собой рудименты хромосом древних бактерий, которые проникли в цитоплазму хозяйских клеток и стали предшественниками этих органелл. Митохондриальная ДНК кодирует митохондриальные тРНК и рРНК, а также несколько митохондриальных белков. Более 95% митохондриальных белков кодируется ядерной ДНК.
СТРОЕНИЕ ГЕНОВ
Рассмотрим строение гена у прокариот и эукариот, их сходства и различия. Несмотря на то, что ген — это участок ДНК, кодирующий всего один белок или РНК, кроме непосредственно кодирующей части, он также включает в себя регуляторные и иные структурные элементы, имеющие разное строение у прокариот и эукариот.
Кодирующая последовательность - основная структурно-функциональная единица гена, именно в ней находятся триплеты нуклеотидов, кодирующие аминокислотную последовательность. Она начинается со старт-кодона и заканчивается стоп-кодоном.
До и после кодирующей последовательности находятся нетранслируемые 5’- и 3’-последовательности . Они выполняют регуляторные и вспомогательные функции, например, обеспечивают посадку рибосомы на и-РНК.
Нетранслируемые и кодирующая последовательности составлют единицу транскрипции - транскрибируемый участок ДНК, то есть участок ДНК, с которого происходит синтез и-РНК.
Терминатор - нетранскрибируемый участок ДНК в конце гена, на котором останавливается синтез РНК.
В начале гена находится регуляторная область , включающая в себя промотор и оператор .
Промотор - последовательность, с которой связывается полимераза в процессе инициации транскрипции. Оператор - это область, с которой могут связываться специальные белки - репрессоры , которые могут уменьшать активность синтеза РНК с этого гена - иначе говоря, уменьшать его экспрессию .
Строение генов у прокариот
Общий план строения генов у прокариот и эукариот не отличается - и те, и другие содержат регуляторную область с промотором и оператором, единицу транскрипции с кодирующей и нетранслируемыми последовательностями и терминатор. Однако организация генов у прокариот и эукариот отличается.

Рис. 18. Схема строения гена у прокариот (бактерий) - изображение увеличивается
В начале и в конце оперона есть единые регуляторные области для нескольких структурных генов. С транскрибируемого участка оперона считывается одна молекула и-РНК, которая содержит несколько кодирующих последовательностей, в каждой из которых есть свой старт- и стоп-кодон. С каждого из таких участков с интезируется один белок. Таким образом, с одной молекулы и-РНК синтезируется несколько молекул белка.
Для прокариот характерно объединение нескольких генов в единую функциональную единицу - оперон . Работу оперона могут регулировать другие гены, которые могут быть заметно удалены от самого оперона - регуляторы . Белок, транслируемый с этого гена называется репрессор . Он связывается с оператором оперона, регулируя экспрессию сразу всех генов, в нем содержащихся.
Для прокариот также характерно явление сопряжения транскрипции и трансляции .
![]()
Рис. 19 Явление сопряжения транскрипции и трансляции у прокариот - изображение увеличивается
Такое сопряжение не встречается у эукариот из-за наличия у них ядерной оболочки, отделяющей цитоплазму, где происходит трансляция, от генетического материала, на котором происходит транскрипция. У прокариот во время синтеза РНК на матрице ДНК с синтезируемой молекулой РНК может сразу связываться рибосома. Таким образом, трансляция начинается еще до завершения транскрипции. Более того, с одной молекулой РНК может одновременно связываться несколько рибосом, синтезируя сразу несколько молекул одного белка.
Строение генов у эукариот
Гены и хромосомы эукариот очень сложно организованы
У бактерий многих видов всего одна хромосома, и почти во всех случаях в каждой хромосоме присутствует по одной копии каждого гена. Лишь немногие гены, например гены рРНК, содержатся в нескольких копиях. Гены и регуляторные последовательности составляют практически весь геном прокариот. Более того, почти каждый ген строго соответствует аминокислотной последовательности (или последовательности РНК), которую он кодирует (рис. 14).
Структурная и функциональная организация генов эукариот гораздо сложнее. Исследование хромосом эукариот, а позднее секвенирование полных последовательностей геномов эукариот принесло много сюрпризов. Многие, если не большинство, генов эукариот обладают интересной особенностью: их нуклеотидные последовательности содержат один или несколько участков ДНК, в которых не кодируется аминокислотная последовательность полипептидного продукта. Такие нетранслируемые вставки нарушают прямое соответствие между нуклеотидной последовательностью гена и аминокислотной последовательностью кодируемого полипептида. Эти нетранслируемые сегменты в составе генов называют интронами , или встроенными последовательностями , а кодирующие сегменты — экзонами . У прокариот лишь немногие гены содержат интроны.
Итак, у эукариот практически не встречается объединение генов в опероны, и кодирующая последовательность гена эукариот чаще всего разделена на транслируемые участки - экзоны , и нетранслируемые участки - интроны.
В большинстве случаев функция интронов не установлена. В целом, лишь около 1,5% ДНК человека являются ≪кодирующими≫, т. е. несут информацию о белках или РНК. Однако с учетом крупных интронов получается, что ДНК человека на 30% состоит из генов. Поскольку гены составляют относительно небольшую долю в геноме человека, значительная часть ДНК остается неучтенной.

Рис. 16. Схема строение гена у эукариот - изображение увеличивается
С каждого гена сначала синтезируется незрелая, или пре-РНК, которая содержит в себе как интроны, так и экзоны.
После этого проходит процесс сплайсинга, в результате которого интронные участки вырезаются, и образуется зрелая иРНК, с которой может быть синтезирован белок.

Рис. 20. Процесс альтернативного сплайсинга - изображение увеличивается
Такая организация генов позволяет, например, осуществить , когда с одного гена могут быть синтезированы разные формы белка, за счет того, что в процессе сплайсинга экзоны могут сшиваться в разных последовательностях.

Рис. 21. Отличия в строении генов прокариот и эукариот - изображение увеличивается
МУТАЦИИ И МУТАГЕНЕЗ
Мутацией называется стойкое изменение генотипа, то есть изменение нуклеотидной последовательности.
Процесс, который приводит к возникновению мутаций называется мутагенезом , а организм, все клетки которого несут одну и ту же мутацию — мутантом .
Мутационная теория была впервые сформулирована Гуго де Фризом в 1903 году. Современный ее вариант включает в себя следующие положения:
1. Мутации возникают внезапно, скачкообразно.
2. Мутации передаются из поколения в поколение.
3. Мутации могут быть полезными, вредными или нейтральными, доминантными или рецессивными.
4. Вероятность обнаружения мутаций зависит от числа исследованных особей.
5. Сходные мутации могут возникать повторно.
6. Мутации не направленны.
Мутации могут возникать под действием различных факторов. Различают мутации, возникшие под действием мутагенных воздействий : физических (например, ультрафиолета или радиации), химических (например, колхицина или активных форм кислорода) и биологических (например, вирусов). Также мутации могут быть вызваны ошибками репликации .
В зависимости от условий появления мутации подразделяют на спонтанные — то есть мутации, возникшие в нормальных условиях, и индуцированые — то есть мутации, которые возникли при особых условиях.
Мутации могут возникать не только в ядерной ДНК, но и, например, в ДНК митохондрий или пластид. Соответственно, мы можем выделять ядерные и цитоплазматические мутации.
В результате возникновения мутаций часто могут появляться новые аллели. Если мутантный аллель подавляет действие нормального, мутация называется доминантной . Если нормальный аллель подавляет мутантный, такая мутация называется рецессивной . Большинство мутаций, приводящих к возникновению новых аллелей являются рецессивными.
По эффекту выделяют мутации адаптивные , приводящие к повышению приспособленности организма к среде, нейтральные , не влияющие на выживаемость, вредные , понижающие приспособленность организмов к условиям среды и летальные , приводящие к смерти организма на ранних стадиях развития.
По последствиям выделяются мутации, приводящие к потери функции белка , мутации, приводящие к возникновению у белка новой функции , а также мутации, которые изменяют дозу гена , и, соответственно, дозу белка синтезируемого с него.
Мутация может возникнуть к любой клетке организма. Если мутация возникает в половой клетке, она называется герминативной (герминальной, или генеративной). Такие мутации не проявляются у того организма, у которого они появились, но приводят к появлению мутантов в потомстве и передаются по наследству, поэтому они важны для генетики и эволюции. Если мутация возникает в любой другой клетке, она называется соматической . Такая мутация может в той или иной степени проявляться у того организма, у которого она возникла, например, приводить к образованию раковых опухолей. Однако такая мутация не передается по наследству и не влияет на потомков.
Мутации могут затрагивать разные по размеру участки генома. Выделяют генные , хромосомные и геномные мутации.
Генные мутации
Мутации, которые возникают в масштабе меньшем, чем один ген, называются генными , или точечными (точковыми) . Такие мутации приводят к изменению одного и нескольких нуклеотидов в последовательности. Среди генных мутаций выделяют замены , приводящие к замене одного нуклеотида на другой, делеции , приводящие к выпадению одного из нуклеотидов, инсерции , приводящие к добавлению лишнего нуклеотида в последовательность.

Рис. 23. Генные (точечные) мутации
По механизму воздействия на белок, генные мутации делят на: синонимичные , которые (в результате вырожденности генетического кода) не приводят к изменению аминокислотного состава белкового продукта, миссенс-мутации , которые приводят к замене одной аминокислоты на другую и могут влиять на структуру синтезируемого белка, хотя часто они оказываются незначительными, нонсенс-мутации , приводящие к замене кодирующего кодона на стоп-кодон, мутации, приводящие к нарушению сплайсинга:

Рис. 24. Схемы мутаций
Также по механизму воздействия на белок выделяют мутации, приводящие к сдвигу рамки
считывания
, например, инсерции и делеции. Такие мутации, как и нонсенс-мутации, хоть и возникают в одной точке гена, часто воздействуют на всю структуру белка, что может привести к полному изменению его структуры.

Рис. 29. Хромосома до и после дупликации
Геномные мутации
Наконец, геномные мутации затрагивают весь геном целиком, то есть меняется количество хромосом. Выделяют полиплоидии — увеличение плоидности клетки, и анеуплоидии, то есть изменение количества хромосом, например, трисомии (наличие у одной из хромосом дополнительного гомолога) и моносомии (отсутствие у хромосомы гомолога).
Видео по теме ДНК
РЕПЛИКАЦИЯ ДНК, КОДИРОВАНИЕ РНК, СИНТЕЗ БЕЛКА
В зависимости от того, какой моносахарид содержится в структурном звене полинуклеотида - рибоза или 2-дезоксирибоза , различают
- рибонуклеиновые кислоты (РНК) и
- дезоксирибонуклеиновые кислоты (ДНК).
Нуклеотидные звенья макромолекул ДНК могут содержать аденин, гуанин, цитозин и тимин . Состав РНК отличается тем, что вместо тимина присутствует урацил .
Молекулярная масса ДНК достигает десятков миллионов а.е.м. Это самые длинные из известных макромолекул. Значительно меньше молекулярная масса РНК (от нескольких сотен до десятков тысяч). ДНК содержатся в основном в ядрах клеток, РНК в рибосомах и протоплазме клеток.
При описании строения нуклеиновых кислот учитывают различные уровни организации макромолекул: первичную и вторичную структуру.
- Первичная структура нуклеиновых кислот это нуклеотидный состав и определенная последовательность нуклеотидных звеньев в полимерной цепи.
В сокращённом однобуквенном обозначении эта структура записывается как ...– А – Г – Ц –...
- Под вторичной структурой нуклеиновых кислот понимают пространственно упорядоченные формы полинуклеотидных цепей.

Такая пространственная структура удерживается множеством водородных связей, образуемых азотистыми основаниями, направленными внутрь спирали. Водородные связи возникают между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи. Эти основания составляют комплементарные пары (от лат. complementum - дополнение). Образование водородных связей между комплементарными парами оснований обусловлено их пространственным соответствием. Пиримидиновое основание комплементарно пуриновому основанию:

Водородные связи между другими парами оснований не позволяют им разместиться в структуре двойной спирали. Таким образом,
- ТИМИН (Т) комплементарен АДЕНИНУ (А),
- ЦИТОЗИН (Ц) комплементарен ГУАНИНУ (Г).

Комплементарность полинуклеотидных цепей служит химической основой главной функции ДНК хранения и передачи наследственных признаков.
Способность ДНК не только хранить, но и использовать генетическую информацию определяется следующими ее свойствами:
Вторичная структура РНК . В отличие от ДНК, молекулы РНК состоят из одной полинуклеотидной цепи и не имеют строго определенной пространственной формы (вторичная структура РНК зависит от их биологических функций).
Основная роль РНК непосредственное участие в биосинтезе белка. Известны три вида клеточных РНК, которые отличаются по местоположению в клетке, составу, размерам и свойствам, определяющим их специфическую роль в образовании белковых макромолекул:
- информационные (матричные) РНК передают закодированную в ДНК информацию о структуре белка от ядра клетки к рибосомам, где и осуществляется синтез белка;
- транспортные РНК собирают аминокислоты в цитоплазме клетки и переносят их в рибосому; молекулы РНК этого типа "узнают" по соответствующим участкам цепи информационной РНК, какие аминокислоты должны участвовать в синтезе белка;
- рибосомные РНК обеспечивают синтез белка определенного строения, считывая информацию с информационной (матричной) РНК.
Относится к нуклеиновым кислотам. Молекулы-полимеры РНК намного меньше, чем у ДНК. Однако в зависимости от типа РНК количество входящих в них нуклеотидов-мономеров различается.
В состав нуклеотида РНК в качестве сахара входит рибоза, в качестве азотистого основания - аденит, гуанин, урацил, цитозин. Урацил по строению и химическим свойствам близок к тимину, который обычен для ДНК. В зрелых молекулах РНК многие азотистые основания модифицированы, поэтому в реальности разновидностей азотистых оснований в составе РНК намного больше.
Рибоза в отличие от дезоксирибозы имеет дополнительную -ОН-группу (гидроксильную). Это обстоятельство позволяет РНК легче вступать в химические реакции.
Главной функцией РНК в клетках живых организмов можно назвать реализацию генетической информации. Именно благодаря разным типам рибонуклеиновой кислоты генетический код считывается (транскрибируется) с ДНК, после чего на его основе синтезируются полипептиды (происходит трансляция). Итак, если ДНК в основном отвечает за хранение и передачу из поколения в поколение генетической информации (основной процесс – репликация), то РНК реализует эту информацию (процессы транскрипции и трансляции). При этом транскрипция происходит на ДНК, так что этот процесс относится к обоим типам нуклеиновых кислот и тогда с этой точки зрения можно сказать, что и ДНК отвечает за реализацию генетической информации.
При более подробном рассмотрении функции РНК намного разнообразнее. Ряд молекул РНК выполняют структурную, каталитическую и другие функции.
Существует так называемая гипотеза РНК-мира, согласно которой вначале в живой природе в качестве носителя генетической информации выступали только молекулы РНК, при этом другие молекулы РНК катализировали различные реакции. Данная гипотеза подтверждена рядом опытов, показывающих возможную эволюцию РНК. На это указывает и то, что ряд вирусов в качестве нуклеиновой кислоты, хранящей генетическую информацию, имеют молекулу РНК.
Согласно гипотезе РНК-мира ДНК появилась позже в процессе естественного отбора как более устойчивая молекула, что важно для хранения генетической информации.
Выделяют три основных типа РНК (кроме них есть и другие): матричная (она же информационная), рибосомальная и транспортная. Обозначаются они соответственно иРНК (или мРНК), рРНК, тРНК.
Информационная РНК (иРНК)
Почти все РНК синтезируются на ДНК в процессе транскрипции. Однако часто транскрипция упоминается как синтез именно информационной РНК (иРНК). Связано это с тем, что последовательность нуклеотидов иРНК в последствии определит последовательность аминокислот синтезируемого в процессе трансляции белка.
Перед транскрипцией нити ДНК расплетаются, и на одной из них с помощью комплекса белков-ферментов синтезируется РНК по принципу комплементарности, так же как это происходит при репликации ДНК. Только напротив аденина ДНК к молекуле РНК присоединяется нуклеотид, содержащий урацил, а не тимин.
На самом деле на ДНК синтезируется не готовая информационная РНК, а ее предшественник - пре-иРНК. Предшественник содержит участки последовательности нуклеотидов, которые не кодируют белок и которые после синтеза пре-иРНК вырезаются при участии малых ядерных и ядрышковых РНК («дополнительные» типы РНК). Эти удаляющиеся участки называются интронами . Остающиеся части иРНК называются экзонами . После удаления интронов экзоны сшиваются между собой. Процесс удаления интронов и сшивания экзонов называется сплайсингом . Усложняющей жизнь особенностью является то, что можно вырезать интроны по-разному, в результате получатся разные готовые иРНК, которые будут служить матрицами для разных белков. Таким образом, вроде бы один ген ДНК может играть роль нескольких генов.
Следует отметить, что у прокариотических организмов сплайсинга не происходит. Обычно их иРНК сразу после синтеза на ДНК готова к трансляции. Бывает, что пока конец молекулы иРНК еще транскрибируется, на ее начале уже сидят рибосомы, синтезирующие белок.
После того как пре-иРНК созревает в информационную РНК и оказывается вне ядра, она становится матрицей для синтеза полипептида. При этом на нее «насаживаются» рибосомы (не сразу, какая-то оказывается первой, другая - второй и т. д.). Каждая синтезирует свою копию белка, т. е. на одной молекуле РНК могут синтезироваться сразу несколько одинаковых белковых молекул (понятно, что каждая будет находиться на своей стадии синтеза).
Рибосома, передвигаясь от начала иРНК к ее концу, считывает по три нуклеотида (хотя вмещает шесть, т. е. два кодона) и присоединяет соответствующую транспортную РНК (имеющую соответствующий кодону антикодон), к которой присоединена соответствующая аминокислота. После этого с помощью активного центра рибосомы ранее синтезированная часть полипептида, соединенная с предшествующей тРНК, как-бы «пересаживается» (образуется пептидная связь) на аминокислоту, прикрепленную к только что пришедшей тРНК. Таким образом, молекула белка постепенно увеличивается.
Когда молекула информационной РНК становится не нужна, клетка ее разрушает.
Транспортная РНК (тРНК)
Транспортная РНК - это достаточно маленькая (по меркам полимеров) молекула (количество нуклеотидов бывает разным, в среднем около 80-ти), во вторичной структуре имеет форму клеверного листа, в третичной сворачивается в нечто подобное букве Г.

Функция тРНК - присоединение к себе соответствующей своему антикодону аминокислоты. В дальнейшем соединение с рибосомой, находящейся на соответствующем антикодону кодоне иРНК, и «передача» этой аминокислоты. Обобщая, можно сказать, что транспортная РНК переносит (на то она и транспортная) аминокислоты к месту синтеза белка.
Живая природа на Земле использует всего около 20-ти аминокислот для синтеза различных белковых молекул (на самом деле аминокислот куда больше). Но поскольку, согласно генетическому коду, кодонов больше 60-ти, то каждой аминокислоте может соответствовать несколько кодонов (на самом деле какой-то больше, какой-то меньше). Таким образом, разновидностей тРНК больше 20, при этом разные транспортные РНК переносят одинаковые аминокислоты. (Но и тут не так все просто.)
Рибосомная РНК (рРНК)
Рибосомную РНК часто также называют рибосомальной РНК. Это одно и то же.
Рибосомная РНК составляет около 80% всей РНК клетки, так как входит в состав рибосом, коих в клетке бывает достаточно много.
В рибосомах рРНК образует комплексы с белками, выполняет структурную и каталитическую функции.
В состав рибосомы входят несколько разных молекул рРНК, отличающиеся между собой как по длине цепи, вторичной и третичной структуре, выполняемым функциям. Однако их суммарная функция - это реализация процесса трансляции. При этом молекулы рРНК считывают информацию с иРНК и катализируют образование пептидной связи между аминокислотами.